Toll-like receptors belong to a family of pattern recognition receptors, regulate innate immunity and subsequent adaptive immune responses. TLRs can up-regulate costimulatory molecules on antigen-presenting cells (APCs) to activate adaptive immune responses. Specific TLR agonists possess high potency and low toxicity to be used as vaccine adjuvants. Furthermore, the TLR1/2 agonists have the potential for chronic and acute inflammatory/infectious disease treatment.
In this article, we will introduce a novel and specific TLR1/2 agonist CU-T12-9 (EC50=52.9 nM). It selectively activates the TLR1/2 heterodimer, not TLR2/6.
As a pivotal part of innate immunity, TLRs can recognize various molecules of microbial origin. At first of all, in HEK-Blue cells, CU-T12-9 significantly activates the SEAP signaling in cells overexpressing hTLR2. However, it does not has an effect on other TLR-overexpressing cells, including TLR3, TLR4, TLR5, TLR7, and TLR8. CU-T12-9 can specifically activate the TLR2 signaling pathway.
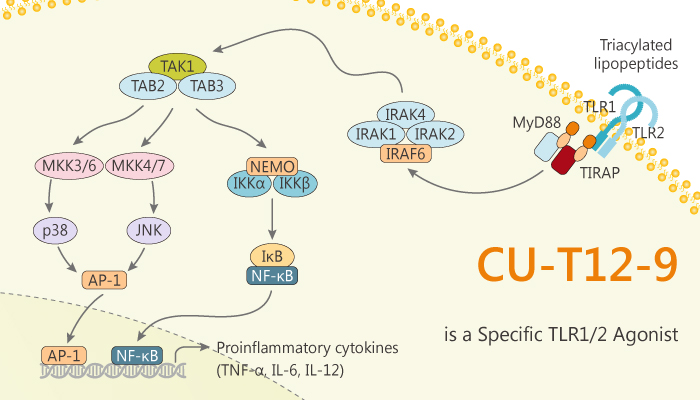
Nextly, as we all know, most TLRs act alone, only TLR2 plays its role by heterodimer with either TLR1 or TLR6.
HEK-Blue hTLR2 cells endogenously express TLR1 and TLR6. In a selectivity experiment with antibody inhibition, the anti-hTLR1 or the anti-hTLR2 antibodies can dose-dependently reverse CU-T12-9-activated SEAP signaling.
By contrast, any inhibition of SEAP signaling cannot be observed by an anti-hTLR6 antibody even its concentrations up to 10 μg/ml. Furthermore, a well-known TLR2/6-specific agonist, Pam2CSK4 is chosen to be a positive control. The result of Pam2CSK4 shows that both anti-hTLR2 and anti-hTLR6 antibodies can inhibit Pam2CSK4-induced SEAP signaling. However, anti-hTLR1 does not influence SEAP expression. These results prove that CU-T12-9 acts by specific activation of TLR1/2 signaling, not TLR2/6.
Lastly, TLRs recruit a set of adaptor proteins through homotypic interactions to trigger downstream signaling cascades. Thereby, it leads to the activation of transcription factor NF-κB.
In aTLR2-sensitive U937 human macrophage cell line with a GFP-labeled NF-κB reporter, CU-T12-9 activates NF-κB signaling in a dose-dependent manner in Flow cytometry experiments.
In addition, CU-T12-9, at 5 μM, shows comparable activation to Pam3CSK4 (66 nM). A known NF-κB inhibitor, triptolide can efficiently inhibit the CU-T12-9–induced SEAP signaling.
Another downstream product of NF-κB activation is nitric oxide (NO). CU-T12-9 significantly triggers NO production in Raw 264.7 cells and also in primary rat macrophage cells.
Furthermore, the TLR1/2-specific antagonist (CU-CPT22) can make an inhibitory effect in NO activation but the TLR4-specific inhibitor TAK242 cannot. All those results further imply that CU-T12-9 works through the NF-κB signaling pathway specifically.
In summary, we can conclude that CU-T12-9, as a novel small-molecule TLR1/2 agonist, recognizes and stabilizes the heterodimer of TLR1/2. It specifically activates TLR1/2 and has potential biomedical applications.
Reference
Cheng K, et al. Sci Adv. 2015;1(3). pii: e1400139.